Биогенетический закон Геккеля-Мюллера описывает наблюдаемое в живой природе соотношение - онтогенез, то есть персональное развитие каждого живого организма, в определенной степени оно повторяет филогенез - историческое развитие всей группы особей, к которой он относится. Закон сформулировали, как понятно из названия, и Ф. Мюллер в 60-е годы XIX века независимо друг от друга, и установить первооткрывателя теории сейчас почти невозможно.
Очевидно, что биогенетический закон не был сформулирован сразу. Работе Мюллера и Геккеля предшествовало создание теоретической базы для закона в виде уже обнаруженных явлений и других установленных закономерностей природы. В 1828 году К. Бэр сформулировал так называемый закон зародышевого сходства. Суть его заключается в том, что эмбрионы особей, относящихся к одному биологическому типу, имеют множество схожих элементов анатомического строения. У человека, например, на определенной стадии развития у зародыша имеются жаберные щели и хвост. Характерные отличительные черты в морфологии видов возникают только в ходе дальнейшего онтогенеза. Закон зародышевого сходства во многом определил биогенетический закон: раз эмбрионы различных организмов повторяют стадии развития других особей, они повторяют стадии развития всего типа вообще.
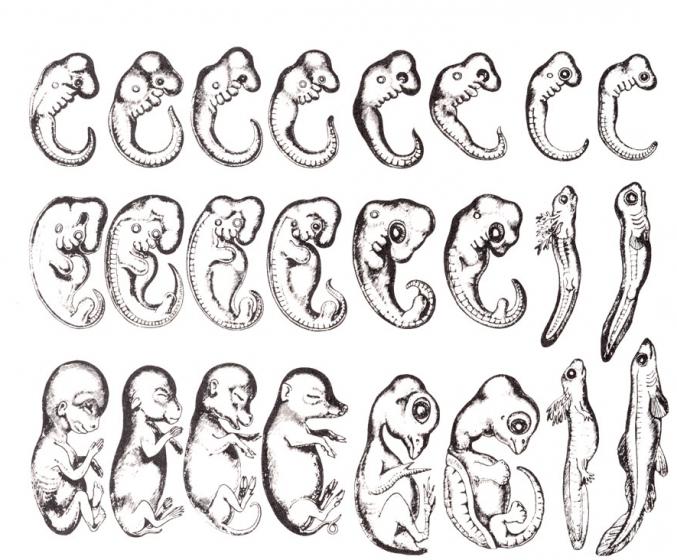
А.Н. Северцовым позднее были внесены определенные поправки к закону Геккеля-Мюллера. Ученый отметил, что во время эмбриогенеза, то есть стадии наблюдается сходство между органами именно эмбрионов, а не взрослых особей. Так, жаберные щели у схожи с жаберными щелями эмбрионов рыб, но никак не со сформировавшимися жабрами взрослых рыб.
Важно отметить, что одним из наиболее значимых доказательств дарвиновской теории эволюции считается непосредственно биогенетический закон. Формулировка его сама по себе намекает на собственную логическую связь с учением Дарвина. Эмбрион в ходе своего развития проходит через множество различных стадий, каждая из которых напоминает определенные ступени развития природы, отмечаемые с эволюционной точки зрения. Таким образом, каждая все более сложно организованная особь отражает в своем онтогенезе развитие всей живой природы с точки зрения эволюции.

В психологии также существует свой биогенетический закон, сформулированный независимо от биологического. По сути, в психологии выделяется не оформленный закон, а высказанная И. Гербартом и Т. Циллером идея о схожести развития психики ребенка с таковой у человечества вообще. Различные ученые пытались обосновать эту теорию с разных точек зрения. Г. Холл, например, прибегал непосредственно к закону Геккеля-Мюллера. Он говорил о том, что развитие ребенка, в том числе в психологическом плане, задается исключительно биологическими предпосылками и повторяет эволюционное развитие вообще. Так или иначе, на сегодняшний день идея не является однозначно доказанной. В психологии до сих пор не существует биогенетического закона как такового.
от греч. ???? – жизнь и??????? – происхождение) – принимаемое эволюционистами-дарвинистами положение, согласно к-рому каждая органич. форма в своем индивидуальном развитии (онтогенезе) в нек-рой мере повторяет черты и особенности тех форм, от к-рых она произошла. Термин "Б. з." введен нем. биологом Э. Геккелем (1866), к-рый сформулировал "основной Б. з.": онтогенез, или развитие органич. индивидуумов как ряд изменений форм, через к-рые проходит каждый индивидуальный организм за время своего индивидуального существования, непосредственно обусловливается филогенезом, т. е. развитием органич. группы, возникшей от общего корня, к к-рому данный организм относится. По Геккелю, "онтогения представляет собой краткое и быстрое повторение (р е к а п и т у л я ц и ю) филогении, повторение, обусловленное физиологическими функциями н а с л е д с т в е н н о с т и (воспроизведения) и п р и с п о с о б л я е м о с т и (питания). В течение быстрого и краткого хода своего онтогенетического развития особь повторяет важнейшие из тех изменений формы, через которые прошли ее предки в течение медленного и длительного хода их палеонтологического развития по законам наследственности и приспособления" (Мюллер Ф. и Геккель Э., Основной биогенетический закон, М.–Л., 1940, с. 169). Геккель различал в индивидуальном развитии п а л и н г е н е з ы, к к-рым относил явления, соответствующие Б. з. и повторяющие те или иные особенности предковых форм, т. е. явления рекапитуляции (повторения), и ц е н о г е н е з ы, к-рые представляют нарушения рекапитуляции, т. е. отступление от повторения филогенеза. На основе палингенезов Геккель считал возможным восстанавливать некогда существовавшие формы, от к-рых происходит данный вид. Б. з. в трактовке Геккеля был грубым упрощением биологич. фактов, хотя и содержал рациональное зерно. На основании данных биологии и эволюционной палеонтологии, или палеобиологии, можно говорить лишь об элементах рекапитуляции в индивидуальном развитии организмов; ни в одной стадии онтогенеза организм не представляет сколько-нибудь полного повторения предковых форм данного филогенетич. ряда. Основная идея Б. з. существовала задолго до Геккеля. О подобии между ступенями зародышевого развития организмов и классификационными рядами менее высокоорганизованных групп писали еще в конце 18 в. и в начале 19 в. как авторы, не имевшие ясного представления об эволюции органич. мира или придерживавшиеся весьма ограниченного понимания эволюции (напр., нем. натурфилософы Л. Окен и И. Меккель), так и нек-рые эволюционисты додарвиновских времен (напр., русский естествоиспытатель К. Ф. Рулье). Впервые проблему соотношения между онтогенезом и филогенезом, проблему рекапитуляции во всей ее полноте поставил Ч. Дарвин. Еще в "Очерке 1844 года", разрабатывая эволюц. теорию, Дарвин писал, что "зародыши ныне существующих позвоночных отражают строение некоторых взрослых форм этого большого класса, существовавших в более ранние периоды истории земли" (Соч., т. 3, 1939, с. 215). В "Происхождении видов..." основная идея теории рекапитуляции и Б. з. выражена Дарвином как закон сходства древних форм с эмбриональными стадиями новейших форм. Дарвин указал многие факты, свидетельствующие в пользу Б. з. (напр., наличие у зародышей млекопитающих и птиц жаберных щелей, у теленка – никогда не прорезывающихся зубов в верхней челюсти там, где у взрослого зубов нет и где они функционально замещаются другими органами). Однако Дарвин отмечал и такие факты, к-рые не укладываются в примитивное понимание Б. з., напр. явления гетерохронии (изменения во времени появления признаков) и, в частности, случаи, когда определенные признаки возникают в онтогенезе потомков раньше, чем в онтогенезе предковых форм. Изменения признаков могут иметь место, по Дарвину, на всех стадиях (яйцо, семя, молодой и взрослый организм); это он объяснял естественным отбором. Явления рекапитуляции рассматривал с дарвинистских позиций нем. ученый Ф. Мюллер (1864). Ф. Энгельс подчеркнул важное значение Б. з. для эволюционного учения. Основная причина явлений рекапитуляции, подчиняющихся Б. з., заключается в том, что эволюция – не внезапная и одновременная полная перестройка всего организма, а процесс последовательного изменения с сохранением, до нек-рого историч. момента, многих основных черт, процесс, к-рый может привести к коренному преобразованию всего тела лишь через многочисленные относительно мелкие изменения, осуществление к-рых требует значительного времени. Многие черты строения и деятельности тела сохраняются на протяжении длинных филогенетич. рядов в виде рудиментарных органов или удерживаются только у юных особей; в обоих случаях они представляют собой прямое доказательство эволюции и того конкретного пути, к-рый она совершала в данной группе. Убедительные примеры рекапитуляции дает изучение многих групп ископаемых животных – плеченогих, пластинчатожаберных и брюхоногих моллюсков, фораминифер и особенно аммоноидей (из класса головоногих моллюсков); в последнем случае рекапитуляция связана с тем, что молодь была близка к взрослым особям по образу жизни. Явления, соответствующие Б. з., имеют значение для систематики организмов, напр. в систематике животных для установления таксономич. положения нек-рых групп низших ракообразных (ракушковых, усоногих); эти явления отмечались в многочисленных группах животных (напр., рудиментарные коренные зубы у юных утконосов, впоследствии совершенно исчезающие; многочисленные зубы у зародышей беззубых китов, затем полностью атрофирующиеся). Рекапитуляция и тесно связанные с ней явления известны не только у животных, но и у растений, на что неоднократно указывал, напр., И. В. Мичурин. Так, рассматривая "влияние атавизма", он отмечал, что оно "является неустранимым во всех без исключения сеянцах различных видов и разновидностей растений только в начальных стадиях развития их из семени, выражаясь тем, что все они в молодости имеют в своем наружном виде кажущееся сходство с дикими формами родоначального вида" (Соч., т. 1, 1948, 185–86). Нек-рые зоологи и палеонтологи совершенно отвергают Б. з., ссылаясь на то, что онтогенез никогда не представляет собой даже краткого повторения важнейших этапов филогенеза. Это возражение имеет силу по отношению к первоначальному геккелевскому определению "основного Б. з.", но не может быть выдвинуто против дарвиновского понимания теории рекапитуляции, согласно к-рому онтогенез отнюдь не представляет краткого повторения филогенеза, но на разных стадиях содержит элементы такого повторения. Столь же неосновательно то возражение против основной идеи Б. з., к-рое основывается на случаях, когда эмбриональные структуры, бесспорно соответствующие некогда существовавшим органам предков данной формы, несут какие-то определенные функции у зародышей, т. е. не являются полностью бесполезными: напр., появление в онтогенезе рыб жаберных щелей, обслуживающих у зародышей дыхательную систему, но в дальнейшем исчезающих, надо рассматривать как рекапитуляцию, так как эти щели соответствуют жаберным щелям предков современных рыб; зубы у зародышей беззубых китов имеют определенное значение в индивидуальном развитии этих животных, но это не опровергает того положения, что такие эмбриональные зубы соответствуют зубам предков, у к-рых они служили для захвата пищи. Нек-рые противники теории рекапитуляции утверждают, что в животном мире господствующее положение занимают процессы, как бы противоположные Б. з. Эти ученые утверждают, что эмбриональные структуры предков часто становятся структурами взрослых особей у потомков – у более поздних представителей данного филогенетич. ряда (нем. палеонтолог О. Шиндевольф, англ. зоолог-эмбриолог Г. де Бир). Крайние представители этого направления приходят к мысли, что эволюция шла в основном от высшего состояния к низшему. Такое понимание соотношения между онтогенезом и филогенезом направлено к дискредитации научной, материалистической эволюционной теории в целом. Лит.: Энгельс Ф., Анти-Дюринг, М., 1957, с. 70; Дарвин Ч., Происхождение видов путем естественного отбора, Соч., т. 3, М.–Л.,1939,с. 627–36; Мичурин И. В., Принципы и методы работы, Соч., Т.1, 2 изд., М., 1948, с. 651; Мюллер Ф., Геккель Э., Основной биогенетический закон, Избр. работы, М.–Л. 1940. Л. Давиташвили. Тбилиси.
Наблюдение двух независимых биологов за онтогенезом организмов позволило сформировать биогенетический закон Геккеля-Мюллера. Впервые формулировка прозвучала в 1866 году. Однако предпосылки становления закона были выявлены ещё в 1820-х годах.
Закон и его значение
Суть закона заключается в том, что в процессе онтогенеза (индивидуального развития организма) особь повторяет формы своих предков и от зачатия до становления проходит стадии филогенеза (исторического развития организмов).
Формулировка зоолога Фрица Мюллера была дана в книге «За Дарвина» в 1864 году. Мюллер писал, что историческое развитие вида отражается в истории индивидуального развития.
Через два года естествоиспытатель Эрнст Геккель сформулировал закон более кратко: онтогенез - быстрое повторение филогенеза. Другими словами, каждый организм проходит эволюционное изменение вида в процессе развития.

Рис. 1. Геккель и Мюллер.
Свои выводы учёные сделали при изучении эмбрионов разных видов на основе ряда схожих признаков. Например, у зародышей млекопитающих и рыб формируются жаберные дуги. Эмбрионы амфибий, рептилий и млекопитающих проходят одинаковые стадии развития и внешне похожи. Схожесть эмбрионов является одним из доказательств теории эволюции и происхождения животных от одного предка.
ТОП-4 статьи которые читают вместе с этой

Рис. 2. Сравнение зародышей разных животных.
Основатель эмбриологии Карл Бэр ещё в 1828 году выявил сходство зародышей разных видов. Он писал о том, что зародыши идентичны и лишь на определённой стадии эмбриологического развития проявляются признаки рода и вида. Любопытно, что, несмотря на свои наблюдения, Бэр так и не принял теорию эволюции.
Критика
С XIX века выводы Геккеля и Мюллера подвергались критике.
Были выявлены несовершенства основного биогенетического закона:
- особь не повторяет все этапы эволюции и проходит стадии исторического развития в сжатой форме;
- сходство наблюдается не у эмбрионов и взрослых особей, а у двух разных эмбрионов на определённом этапе развития (жабры млекопитающих схожи с жабрами зародышей рыб, а не взрослых особей);
- неотения - явление, при котором взрослая стадия напоминает личиночное развитие предполагаемого предка (сохранение на протяжении всей жизни младенческих свойств);
- педогенез - вид партеногенеза, при котором размножение происходит на стадии личинки;
- значительные различия на стадиях бластулы и гаструлы у позвоночных, сходство наблюдается на более поздних стадиях.
Установлено, что закон Геккеля-Мюллера никогда не выполняется полностью, всегда находятся отклонения и исключения. Некоторые эмбриологии отмечали, что биогенетический закон - всего лишь иллюзия, не имеющая под собой серьёзных предпосылок.
Закон пересмотрел биолог Алексей Северцов. На основе биогенетического закона он разработал теорию филэмбриогенеза. Согласно гипотезе изменения исторического развития обуславливаются изменениями на личиночной или эмбриональной стадии развития, т.е. онтогенез изменяет филогенез.
Северцов разделил признаки эмбрионов на ценогенезы (приспособления к личиночному или эмбриональному образу жизни) и филэмбриогенезы (изменения эмбрионов, которые приводят к видоизменению взрослых особей).
К ценогенезу Северцов относил:
- зародышевые оболочки;
- плаценту;
- яйцевой зуб;
- жабры личинок земноводных;
- органы прикрепления у личинок.

Рис. 3. Яйцевой зуб - пример ценогенеза.
Ценогенез «облегчил» жизнь личинок и эмбрионов в ходе эволюции. Поэтому сложно проследить развитие филогенеза по эмбриологическому развитию.
Филэмбриогенез делится на три вида:
- архаллаксис - изменения на первых стадиях онтогенеза, при котором дальнейшее развитие организма идёт по новому пути;
- анаболия - увеличение онтогенеза путём возникновения дополнительных стадий эмбрионального развития;
- девиация - изменения на средних стадиях развития.
Немецкие ученые Ф. Мюллер (1828) и Э. Геккель (1866) установили закон соотношения онтогенеза и филогенеза, который получил название биогенетического закона. Согласно этому закону, онтогенез всякого организма есть краткое повторение (рекапитуляция) основных этапов филогенеза вида, к которому данный организм принадлежит. Э. Геккель и Ф. Мюллер считали, что последовательные изменения формы особи на протяжении индивидуального развития обусловлены филогенезом, т. е. развитием того рода, к которому принадлежит данный вид животного. Согласно биогенетическому закону, зародыши систематически вышестоящих животных похожи на взрослых нижестоящих предков. Эволюция осуществляется благодаря прибавлению новых стадий в конце развития.
Биогенетический закон находит множество подтверждений в данных сравнительной анатомии, эмбриологии и палеонтологии. Напр., у зародышей птиц и млекопитающих на определённой стадии эмбрионального развития появляются зачатки жаберного аппарата. Это объясняется тем, что наземные позвоночные произошли от дышавших жабрами рыбообразных предков.
Однако за короткий период индивидуального развития особь не может повторить все этапы эволюции, которая совершалась тысячи или миллионы лет. Поэтому повторение стадий исторического развития вида в индивидуальном развитии особи происходит в сжатой форме, с выпадением ряда этапов. Кроме того, эмбрионы имеют сходство не со взрослыми формами предков, а с их зародышами. Так, в онтогенезе млекопитающих имеется этап, на котором у зародышей образуются жаберные дуги. У зародыша рыбы на основании этих дуг образуется орган дыхания - жаберный аппарат. В онтогенезе млекопитающих повторяется не строение жаберного аппарата взрослых рыб, а строение закладок жаберного аппарата зародыша, на основе которых у млекопитающих развиваются совершенно иные органы.
Опираясь на биогенетический закон и используя данные эмбриологии, можно воссоздавать ход исторического развития тех или иных групп организмов. Это особенно важно в тех случаях, когда для какой-либо группы неизвестны ископаемые остатки предковых форм, т. е. при неполноте палеонтологической летописи. Приведем лишь один классический пример. Систематическое положение и происхождение асцидий (Ascidiae ), ведущих сидячий образ жизни, долгое время были совершенно неясны, и только знаменитое исследование А. О. Ковалевского (1866) по развитию этих животных окончательно решило вопрос. Из яйца асцидий выходит свободноплавающая хвостатая личинка, сходная по плану строения с хордовыми (Chordata ). Во время метаморфоза осевшей на дно личинки хвост с хордой и мускулатурой и органы чувств исчезают, нервная трубка редуцируется до степени небольшого нервного узелка, происходит усиленное разрастание брюшной поверхности тела, образуются сифоны и т.д., т.е. появляются особенности организации, связанные с сидячим образом жизни. Сформированная молодая асцидия не имеет уже почти ничего общего с другими хордовыми животными. В этом примере личинка своей организацией рекапитулирует (повторяет) главные черты строения свободноплавающего предка. Так было найдено естественное место асцидий в системе животного царства.
Особенный интерес для эволюционной зоологии представляют рекапитуляции, т.е. повторения в ходе индивидуального развития характерных особенностей строения более или менее отдаленных предков. Типичный пример рекапитуляции приводит академик А.Н. Северцов в своей книге «Морфологические закономерности эволюции», изданной в 1913 г. в Германии. У современных бесхвостых амфибий во взрослом состоянии большая и малая берцовая кости срослись между собой, а у головастиков они разделены. У стегоцефалов, от которых произошли современные амфибии, тоже имелись две отдельные берцовые кости. Следовательно, наличие раздельных берцовых костей у головастиков можно рассматривать как рекапитуляцию одного из признаков задних конечностей, свойственных предковому скелету.
Рекапитуляция не ограничивается морфологическими признаками. Их можно выявить и при онтогенетическом становлении функций различных органов и тканей. Известно, что в процессе эволюции позвоночных постепенно утрачивались ферменты, необходимые для расщепления мочевой кислоты. Так, у некоторых рептилий и птиц конечный продукт азотистого обмена - мочевая кислота, у земноводных и большинства рыб - мочевина, а у первичновидных беспозвоночных - аммиак. Оказалось, что зародыш птиц на ранних стадиях развития выделяет аммиак, на более поздних - мочевину и только на последних стадиях - мочевую кислоту. Сходным образом у головастиков конечный продукт обмена составляет аммиак, а у лягушек - мочевина.
Проявлению в неизменном виде первобытных, древних, палингенетических признаков (от греч. palaios - древний; палингенезы - признаки, перешедшие в онтогенез животного из его филогенеза) мешают ценогенезы - разнообразные признаки, возникшие в онтогенезе как приспособление к условиям жизни зародышей личинок и взрослых животных. Примером ценогенезов могут служить амнион, хорион, аллантоис Amniota, громадные прядильные железы личинок насекомых и др.
Дальнейшие эмбриологические исследования (А.Н. Северцов, И.И. Шмальгаузен) показали, что в теории Мюллера-Геккеля было много упущений, и главное из них заключалось в том, что история взрослого организма рассматривалась в отрыве от истории эмбриона. Несмотря на это, нельзя недооценивать огромное значение биогенетического закона в развитии эволюционного учения.
Современные эмбриологические исследования показали, что закон верен лишь в общих чертах.
Нет ни одной стадии в развитии, на которой зародыш полностью повторял бы строение какого-либо предка по филогенезу;
В онтогенезе повторяется строение не взрослых стадий предков, а их эмбрионов. Например, зародыш млекопитающих никогда целиком не повторяет строение рыбы, но на определенной стадии развития у него закладываются жаберные щели и жаберные артерии.
Биогенетический закон Геккеля – Мюллера или основной биогенетический закон гласит: каждое живое существо в своем индивидуальном развитии (онтогенез) повторяет в известной степени формы, пройденные его предками или его видом (филогенез).
Этот закон сыграл важную роль в истории развития науки, однако в настоящее время в своем исходном виде не признается современной биологической наукой.
По современной трактовке биогенетического закона, предложенного русским биологом А. Н. Северцовым в начале ХХ века, в онтогенезе происходит повторение признаков не взрослых особей, а их предков.
Часто с биогенетическим законом сопоставляют закон зародышевого сходства, сформулированный К. М. Бэром в 1828 году, из которого следует, что эмбрионы последовательно проходят в своем развитии от общих признаков типа ко все более специальным признакам; позднее всего развиваются признаки, указывающее на принадлежность эмбриона к определенному роду, виду, и, наконец, развитие завершается появлением характерных особенностей данной особи.
Ряд исследователей (Северцев, 1939; Шмальгаузен, 1969, Иванова – Казас, 1939) показали, что закон зародышевого сходства и биогенетический закон обуславливаются различными механизмами, и, соответственно, это два разных закона.
Северцев (1939) утверждал, что у одноклеточных онтогенез отсутствует, и только для вольвоксов признавал его наличие в самой примитивной форме.
Вслед за Северцовым большинство эмбриологов отрицает наличие индивидуального развития на клеточном уровне организации, однако, при таком подходе непонятно на базе чего сформовались морфогенетические механизмы первых многоклеточных животных?
Словосочетание «онтогенез клетки», вероятно, первым использовал Бауэр (1935). Ученик Баэра - Токин (1939) обратил внимание, что особь (томит), образованная в результате деление брюхоресничной инфузории, получает разный набор цирр (ресничных пучков) и, соответственно, должна восстановить недостающие пучки. Токин трактовал такой процесс как онтогенез, а последний этап формирования нового ресничного аппарата он считал рекапитуляциями.
Инфузории, в отличие от других протистов, имеют клеточное тело с хорошо заметными, даже на светооптическом уровне, внешними структурами. В первую очередь – это клеточный рот, для инфузории характерно поперечное деление, в результате которого одна из дочерних клеток одна из дочерних клеток получает цистом (и соответствующий ресничный аппарат), а другая должна его достраивать. Как оказалось, построение нового ротового аппарата (стоматогенез) протекает в разных группах инфузорий по – разному. Последовательность процессов, которые протекают в стоматогенезе инфузорий, трактуются как онтогенез.
Этапы стоматогенеза рассматриваются в качестве рекапитуляций.
Корлисс (Corliss, 1968) полагает, что во многих таксонах простейших можно найти примеры рекапитуляций предковых признаков. Особенно это касается групп со сложно сформированными наружными структурами: пелликулой или различного рода скелетными образованиями. По его мнению, такие примеры можно найти у миксоспоридий (скульптура стенок спор), грегарин (мембрана гаметоцист) и т.д.
И. В. Довгаль, в отличие от других протистологов считает, что стоматогенез инфузорий и ранние этапы метаморфоза расселительных стадий сидячих инфузорий (и аналогичные процессы у других групп одноклеточных) – проявление не биогенетического закона, а закона зародышевого сходства Бэра (Довгаль, 2000; Dovgal, 2002). За основу он принимает формулировку закона зародышевого сходства из монографиии И. И. Шмальгаузена (1969).
Биогенетический закон не справедлив для одноклеточных. Для протистов справедлив закон зародышевого сходства Бэра.